|
||||
|
|
Глава V ЛОГИКА НЕРВНОЙ СИСТЕМЫ СТРУКТУРА РЕФЛЕКСОВ – КЛАССИЧЕСКАЯ ТОЧКА ЗРЕНИЯ В главе III было показано, насколько полезен анализ того, как построен и как работает рецептор, для моделирования некоторых сторон организации центральной нервной системы. Продолжая такой анализ, мы обратимся в настоящей главе к рассмотрению выходных механизмов мозга, полагая, что и эти процессы могут быть богатым источником данных о конструкции мозговых механизмов, создающих возможность кодирования. Модели, построенные на основе этих двух источников, весьма сходны, что лишний раз свидетельствует об адекватности принятого нами подхода. При рассмотрении механизмов выходных систем доступность поведения наблюдению позволяет лучше понять формальные характеристики, саму логику нейрофизиологического процесса. Эта логика, нашедшая свое воплощение в классическом понятии рефлекторной дуги, в настоящее время рассматривается как процесс, который выполняет как аналитическую, так и контрольную функции. Чтобы разрешить противоречие между новыми данными и ранее широко распространенной теорией, необходимо остановиться на истории вопроса. В работе Шеррингтона «Интегративная деятельность нервной системы» сформулирована классическая концепция организации рефлексов. Эта концепция не только дала толчок большому числу нейрофизиологических исследований (D. Lloyd, 1959), но оказала даже еще более глубокое влияние на психологическую мысль. Шеррингтон, пытаясь определить единицу анализа поведения «спинального» препарата, который он изучал, рассматривал организацию рефлекса следующим образом. «Возможно, «простой рефлекс» является чисто абстрактным понятием, так как все части нервной системы связаны воедино и, вероятно, ни одна из них не в состоянии участвовать в какой-либо реакции, не воздействуя и не испытывая воздействия со стороны других частей, причем вся система, несомненно, никогда не находится в состоянии полного покоя. Однако понятие «простая рефлекторная реакция» оправданно, хотя и несколько проблематично (Шеррингтон, 1969, стр. 35). Представления Шеррингтона основаны на ранних экспериментах Белла (1811) и Мажанди (1822). Эти исследователи успешно использовали анатомические данные о том, что каждый соматический периферический нерв в месте соединения со спинным мозгом разделяется на две главные ветви: на вентральный и дорсальный корешки. Перерезка дорсальных корешков нервов конечности собаки вызывает анестезию (полную потерю чувствительности) этой конечности без какого-либо заметного изменения характера ее двигательных реакций (моторной функции). Перерезка вентральных корешков нервов конечности вызывает ее паралич без всякого изменения чувствительности. Результаты этих экспериментов были настолько важны, что они были обобщены в закон (закон Белла и Мажанди), который гласит, что входные-и выходные сигналы в нервной системе передаются по изолированным путям. Поэтому входные нервы были названы афферен-тами (ad +fего = приносить), а выходные – эфферентным» (ех +fero = выносить) (см. рис. V-1). 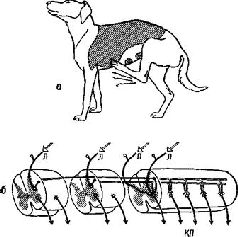
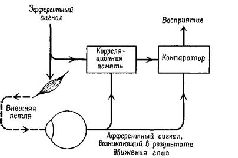
Долгое время из закона Белла и Мажанди делали вывод, что все афференты являются сенсорными (то есть связанными с сенсорными рецепторами) нервами, а все эфференты – моторными (то есть связанными с сокращающейся мускулатурой). Это обобщение получило свое отражение в знаменитом понятии! рефлекторной дуги Шеррингтона. Рефлекторная дуга является нейрологическим выражением бихевиористского подхода к анализу психологических процессов в терминах простого соответствия между входом организма (стимулами) и выходом (ответами). ЭФФЕРЕНТНЫЙ КОНТРОЛЬ ВХОДА – ПЕТЛЯ ОБРАТНОЙ СВЯЗИ Пересмотр концепции рефлекторной дуги стал необходим из-за новых экспериментальных данных, которыми не располагал Шеррингтон. Эти данные показывают, что все входные системы организма непосредственно контролируются центральной нервной системой. Таким образом, выходные волокна – эфференты – регулируют не только движения организма, но и его рецепторы» а следовательно, и его сенсорные функции. Такой пересмотр классического понятия рефлекторной дуги начался с экспериментов, которые продолжали анализ функций нервных волокон, образующих вентральный корешок (см. рис. V-2). 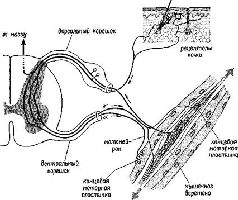
Анатомический анализ показал, что одна треть этих волокон – группа волокон с маленьким диаметром, названная у-волокнами (в отличие от а- и р-волокон – большого диаметра)», оканчивается не в самой ткани сокращающихся мышц, а в особых рецепторах, названных мышечными веретенами. Эти веретена получают информацию о величине напряжения мышечных волокон, внутри которых они находятся. То, что эти веретена» иннервирует такое большое число волокон центрального корешка, весьма озадачило физиологов, разделявших концепцию рефлекторной дуги. Были проведены новые эксперименты, чтобы найти решение этой проблемы. Наиболее поразительные результаты, были получены при электрическом раздражении дистального (обращенного к мышце) конца перерезанного вентрального корешка у которого были удалены путем рассечения волокна большого диаметра. Оказалось, что такая стимуляция не вызывает непосредственно сокращения мышц и, следовательно, у-волокна не являются зфферентами, а их функция – моторной. Еще более интересные результаты были получены во время стимуляции Y-эфферентных волокон и записи электрической активности дорсального корешка. В результате стимуляции вентрального корешка оказалась заметно сниженной спонтанная активность в афферентах дорсального корешка (Granit, 1944). Таким образом, Y-система образует отрицательную обратную связь, регулирующую вход, она берет свое начало от мышечных веретен, действуя в значительной степени так же, как механизмы адаптации и привыкания, описанные в гл. III. Далее было установлено, что активность афферентов мышечных веретен испытывает влияние через у-эфферентные связи со стороны спинного мозга (клеток малого вентрального рога), ствола мозга, мозжечка и даже коры (Kuffler .and Hunt, 1952; Leskell, 1945). Как будет подробно рассмотрено в гл. XII и XIII, эти экспериментальные результаты заставляют предположить, что регуляция моторных функций организма, его-поведения осуществляется скорее посредством особых рецепторов, которые тесно связаны с движением, а не в результате прямого управления мышечными сокращениями. Поэтому мы отложим дальнейшее изложение наших соображений по поводу нейронного контроля поведения до тех пор, пока не проанализируем в деталях тот механизм, с помощью которого осуществляется организация происходящих в рецепторах процессов. Вначале думали, что моторная система уникальна и центральный контроль над рецепторами существует только в двигательной системе. Однако очень скоро стали накапливаться данные о том, что кожные рецепторы (Hagbarth and Kerr, 1954), слуховой афферентный механизм (Galambos, 1956; Rasmussen, 1946; Desmedt r 1960; Dewson, 1968) и обонятельная чувствительность (Kerr and Hagbarth, 1955) также являются объектом прямой регуляции со‹ стороны центральной нервной системы. Не сразу удалось доказать существование эфферентного контроля зрительного входа; первые исследования (Granit, 1955; Hernandez-Peon and Scherer 1955) были подвергнуты критике потому, что еще не найдены морфологически эфферентные волокна в сетчатке. Поэтому в моей лаборатории были проведены исследования в надежде, что эфферентный контроль над зрительным входом станет вполне реальным фактом. В этих исследованиях были использованы только что появившиеся компьютеры и микроэлектродная техника. Результаты опытов показали, что незрительные стимулы (звуковые щелчки и удары по лапе) вызывают ответы в зрительном нерве у кошек (мышцы которых, в том числе и мышцы зрачка, были обездвижены). Кроме того, электрическая активность -сетчатки (измеряемая с помощью электроретинограммы) и аффе-рентов, берущих начало в сетчатке, также меняется в результате такой незрительной стимуляции (Spinelli, Pribram and Weingar-ten, рис. V-3).  Рис. V-3. Запись электрических ответов на звуковой щелчок с помощью биполярных электродов, вживленных в зрительный нерв. Запись осуществлена непосредственно на фотопленку с осциллоскопа методом суперпозиции. Левая запись была сделана, когда животное обнаруживало реакцию внимания, а правая – во время его отвлечения (Spinelli, Pribram and Weingarten, 1965). Наконец, как и в случае других сенсорных модальностей, стимуляция соответствующей части коры головного мозга вызывала изменение в рецептивных полях ганглиозных клеток сетчатки, например их величины (Spinelli and Pribram, 1967). Результаты этих экспериментов убеждают в том, что организация зрительного механизма сходна с организацией других сенсорных систем, в которых существует центральный контроль над сенсорным выходом. МЕХАНИЗМЫ ОПЕРЕЖАЮЩЕГО ВОЗБУЖДЕНИЯ Один процесс, участвующий в нейронном контроле над входом, отличается от до сих пор рассмотренных обратных связей. Интерес к нему возник после того, как великий немецкий физик и психофизиолог Герман Гельмгольц обнаружил некий парадокс в области восприятия. Он заметил, что зрительное изображение мира начинает прыгать, когда мы пальцем надавливаем на наше глазное яблоко. Напротив, при произвольном движении глаз или при движении ими в ответ на внешние стимулы зрительное изображение мира остается неподвижным. В обоих случаях при манипулятивном и обычном движении глаз зрительные структуры, проецирующиеся на сетчатку, идентичны. Гельмгольц сделал вывод, что обычный процесс восприятия должен, следовательно, включать некий механизм, рассчитывающий и корригирующий сигналы, поступающие от сетчатки, насколько это необходимо для получения неискаженного образа. Мертон (Brindley and Merton, 1960) проверил наблюдения Гельмгольца экспериментально. Он парализовал мышцы своего собственного глаза, а затем попытался двигать глазами. Оказалось, что изображение зрительного мира прыгало в том же направлении, в каком он пытался переместить свой взор. Было бы логично предположить, что в осуществлении коррекции, необходимой для обычного восприятия мира, принимает участие обратная связь от мышц глаза. Однако результаты экспериментов с параличом мышц глаза плохо согласуются с этой гипотезой, поскольку движение изображения зрительного мира не связано ни с какими сокращениями мышц. Кроме того, пути, проводящие проприоцептивные импульсы от мышц глаза, не были прослежены в мозгу, несмотря на многочисленные попытки это сделать. Все эти факты заставляют предположить, что существует какой-то «опережающий» процесс, который корректирует восприятие (МасКау, 1966; Mittelstaedt, 1968). 

Неврологические «доказательства», подтверждающие существование опережающего, или, как пазвал его Тейбер (I960), «результативного разряда», были получены недавно. Однако они пока не являются ни прямыми, ни полными. Всякий раз, как возникают движения глаз, на высших уровнях сенсорной системы наблюдаются разряды нейронов (Bizzi, 1966a, 19666, рис. V-4, V-5). Более того, двигательному ответу глаза предшествует появление в зрительной коре определенных динамических структур. По-видимому, каждая из этих динамических структур специфическим образом связана с определенным движением глаз и появляется только после того, как животное (обезьяна) научилось решать задачу, которая требовала выполнения этих двигательных реакций (Pribram, Spinelli and Kamback, 1967). Эти «системы намерения» будут подробно обсуждаться в гл. VII и XVII. ПРОБА – ОПЕРАЦИЯ – ПРОБА – РЕЗУЛЬТАТ (Т – О – Т – Е) Сторонники теории рефлекторной дуги вынуждены считаться со всеми этими фактами. Очевидно самой простой и непосредственной модификацией теории Шеррингтона является признание существования связи между рецептором и выходом центральной нервной системы. Выводы, следующие из этого на первый взгляд небольшого дополнения, отнюдь не тривиальны. Представьте себе, что вы, будучи изолированы от других стимуляций, регистрируете активность своего афферентного нерва. Как вы узнали бы при изменении электрической активности афферентного волокна, являются ли эти изменения результатом воздействия извне или следствием модификации рецепторной активности за счет процессов, возникающих внутри самой центральной нервной системы? Прежде чем ответить на этот вопрос, необходимо произвести некоторые пробы («тестирование»), выполнить некоторые расчеты, чтобы распознать «реальность», то есть действие внешнего стимула (рис. V-6). 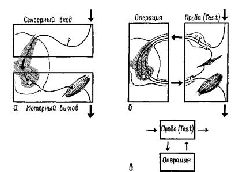
Многочисленные данные, полученные при изучении поведения, подтверждают представление о том, что на входе центральной нервной системы осуществляется некоторый вид подобных активных проб. Многие из относящихся к этой проблеме наблюдений были получены в экспериментах, связанных с искажением зрительного образа или переворачиванием его с помощью специальных очков, которые длительное время носил субъект. Благодаря тому, что организм мог при этом двигаться и манипулировать окружающими предметами, уже через несколько дней или даже часов (в зависимости от степени искажения образа) субъект мог восстановить правильное восприятие мира. Однако такая манипуляция с предметами и движение не могли бы осуществляться, если бы коррекция искажений происходила бы со значительной издержкой, если сама коррекция вообще возможна. Манипуляция предметами является неотъемлемой частью некоторой фазы механизма коррекции (Held, 1968; Howard, Craske and Templeton, 1965; рис. V-7, V-8). 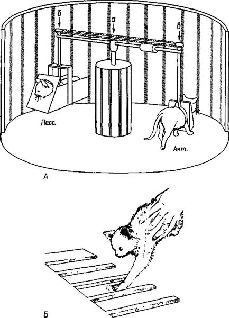
Некоторое представление о силе этого тестирующего механизма можно получить следующим образом. Подойдите к неподвижному эскалатору, который обычно поднимал вас вверх. Сделайте шаг и встаньте на него. Вам сразу же покажется, что ступени несколько секунд движутся вверх. При этом у вас может даже возникнуть желание схватиться за поручень, чтобы удержаться от падения. Это происходит в результате рассогласования ваших соматических ощущений, сигнализирующих об отсутствии движения эскалатора, и зрительного образа кажущегося движения ступенек. Как только вы встали на ступеньки, ощущение движения тут же прекращается. Теперь начните подниматься по ступенькам, и вы снова получите ощущение движения лестницы. «Движение» появляется в результате вмешательства специфической установки, связанной с вашим прошлым опытом в отношении эскалатора (обычные лестницы не воспринимаются как движущиеся). Эта установка усиливается в результате собственного движения по эскалатору. Тот факт, что процесс опробования так сильно зависит от возможности организма манипулировать с окружающей средой, заставляет предположить, что восприятие есть, в сущности, «моторное» явление (Sperry, 1952; Festinger et Ы., 1967). Согласно таким взглядам, восприятие является скорее отражением динамических структур, регулирующих реакции, возникающие в мозгу в ответ на входное воздействие, чем просто результатом самого входного воздействия. Доведенная до своего логического завершения, такая точка зрения привела бы нас к абсурдному заключению. Она означала бы, что мы воспринимаем каждую женщину на улице как идеальный образ Афродиты, а каждого мужчину – как такой же образ Адониса. Конечно, нужно сделать поправку на то, что у зрителя существует свое представление о «красоте». Однако возможные искажения восприятия весьма ограничены. Когда же они выходят за эти границы, мы говорим об иллюзиях и галлюцинациях. И хотя любое объяснение перцептивных процессов должно предполагать возможность появления таких иллюзий и галлюцинаций, оно не должно исходить из того, что иллюзии и галлюцинации являются нормой восприятия. Таким образом, можно попытаться создать обобщенную схему рефлекса, единицу поведенческого анализа (см. Т-О-Т-Е, рис. V-9). Чтобы быть эффективным, входное воздействие должно сравниваться и сопоставляться со спонтанной или завершающей активностью центральной нервной системы. Сигналы, возникающие в результате такого сравнения, вызывают некоторую операцию, оказывающую затем влияние на другие части нервной системы или на внешний мир. Сигналы о результатах такой операции посылаются назад к компаратору. И петля обратной связи продолжает функционировать до тех пор, пока тестирование не даст удовлетворительных результатов, то есть пока не будет достигнуто некоторое ожидаемое состояние (выходной процесс). В результате такой модификации схемы рефлекторной дуги возникает система, хорошо знакомая инженерам. По таким схемам строятся разного рода следящие устройства. Система, известная под именем сервомеханизма, по существу, является устройством, в котором входные воздействия сравниваются с результатами его действий, назначение которых состоит в том, чтобы наменять входные воздействия. Наиболее известным из таких сервомеханизмов является, вероятно, термостат. С помощью своей концепции рефлекторной дуги Шерринг-тон мог объяснить все известные ему экспериментальные данные. Успех такого объяснения способствовал тому, что схема рефлекторной дуги оказалась крайне полезной фикцией (моделью). Схема Т-О-Т-Е также является фикцией, когда она используется для нейропсихологического анализа. Она является в каком-то смысле фикцией более высокого порядка, чем рефлекторная дуга, поскольку рефлекторная дуга – частный случай сервомеханизма , в котором обратная связь может быть реализована только через посредство окружающей среды и выполняемая им операция даже не контролируется зтой обратной связью. Иными словами, действие этого сервомеханизма, однажды начавшись, продолжается до достижения ранее предопределенного состояния. Полезность модели более высокого порядка заключается в ее способности охватить большее число фактов. Именно по этой причине и возникла концепция Т-О-Т-Е, так как теория рефлекторной дуги не может удовлетворительно объяснить данные, которые указывают на существование центрального контроля над рецепторными механизмами. Далее концепция Т-О-Т-Е может объяснить множество других нейропсихологических данных, таких, как процессы адаптации и привыкания, рассмотренные в гл. III. Важно, однако, иметь в виду, что нейропсихологическая концепция Т-О-Т-Е, так же как и теория рефлекторной дуги, япляется только моделью и поэтому она должна быть заменена другой или дополнена, если окажется, что она ограничивает исследование, а не приносит ему пользу. Более детально разработанная, но еще очень упрощенная схема Т-О-Т-Е показана на рис. V-9. По сравнению с предыдущей она яснее показывает отношения между обратной и опережающей связями, а также роль кодирования, памяти и механизма коррекции в нейропове-денческом процессе. 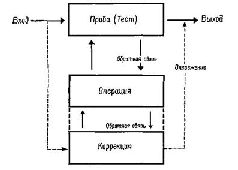
В создании стройной системы взглядов на нейронную организацию поведения концепции Т-О-Т-Е как дальнейшему развитию теории рефлекторной дуги принадлежит центральное место (как пример одной такой последовательно развиваемой точки зрения см. Миллер, Галантер и Прибрам, 1965) (К сожалению, автор не ссылается здесь на концепцию «функциональных систем», «опережающего возбуждения» и «акцептора результатов, действия», детально разработанную в советской физиологии акад. П. К. Анохиным; по-видимому, эта концепция осталась недостаточно известной автору. Прим. ред.) ЗАКЛЮЧЕНИЕ Задача первой части книги состояла в том, чтобы собрать воедино множество разрозненных фактов о работе мозга и его контроле над поведением, чтобы создать какую-то упрощенную схему, которая могла бы служить основой для дальнейших размышлений и исследований. Такой схемой может быть схема Т-О-Т-Е. Ее логический «элемент» состоит из двух реципрокно связанных между собой процессов: 1) «опробования», или «теста», – состояния, представленного потенциалами соединений (синаптическими п дендритными), и 2) «операции», то есть действия, направленного на изменение этого состояния, что реализуется посредством нервных импульсов, генерируемых в рецепторах или в центральной нервной системе. Этот логический элемент, включающий в себя два процесса, является, разумеется, схемой, значительно упрощающей существующие отношения. Но я хотел бы подчеркнуть полезность концепции Т-О-Т-Е для нашего времени и наших исследовательских задач, точно так же, как Шеррингтон подчеркивал полезность теории рефлекторной дуги для своего времени. Блок «состояния» или «коррекции» вводит в схему Т-О-Т-Е. «процесс усиления контраста», создаваемого за счет периферического торможения. Блок «тестирования» (сличения входного воздействия с существующим состоянием) наряду с другими факторами включает процесс пространственного наложения (суперпозиции) возбудительных и тормозных динамических структур взаимодействий, возникающих между соседними нейронами. Операторный блок схемы Т-О-Т-Е Еключает наряду с другими механизмами процесс затухания (ослабления), препятствующий появлению реакций на входные сигналы в каждом нейроне или лейронном пуле. Пространственная суперпозиция увеличивает контраст и таким образом облегчает кодирование; переход в латентное состояние является одной из многих форм сохранения следов памяти (рис. V-10). 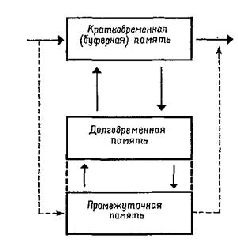
Из-за того, что группы нейронов обладают спонтанной активностью – цикличной или осуществляющейся по определенной программе, – причины изменения состояний в нервной системе заключены не только в окружающей среде, но также и в самом мозгу. Этот факт, а также данные, говорящие о существовании центрального контроля над рецепторной функцией, делают концепцию рефлекторной дуги (стимула – реакции) почти бесполезной для объяснения нервной организации поведения, не говоря уже о психологических функциях. В следующих частях книги будет подробно обоснована необходимость рассматривать все наделенные чувствительностью организмы -и особенно человека – как системы, активно отвечающие на требования окружающей среды. На Западе существовали две основные точки зрения в вопросе об отношении человека к окружающему его миру. Сторонники одной из них утверждают, что человек, в сущности, пассивный организм, сформировавшийся под воздействием окружающей среды. Сторонники другой, напротив, подчеркивают его активную роль, манипулятивный и избирательный характер не только ею ответов, но и чувственных данных. Нейропсихологи-ческий вклад в науку о поведении подтверждает представление об активной природе человека и, таким образом, способствует возрождению звания человека в качестве как научной, так и гуманитарной доктрины. (Pribram, Neuropsychology in America в: В Berelson (ed.). The Behavioral Scionces Today, New York, Basic book, p. 110-111.) РЕЗЮМЕ Гипотеза о двухпроцессном механизме функционирования мозга вытекает из целого ряда данных. Согласно классическому представлению функционирование нервной системы рассматривалось по принципу прямого отношения входа – выхода, стимула – реакции, что получило свое неврологическое выражение в понятии рефлекторной дуги. Однако наличие механизма обратной связи и опережения (то есть контроля со стороны центральной нервной системы за деятельностью рецепторов) вызывает необходимость пересмотра такого представления. Обратная связь и Опережение получили наилучшее обоснование в концепции сервомеханизма Т-О-Т-Е, элементарной логической нейронной структуры, на основе которой создаются более сложные нервные организации. Фаза опробования («теста») выражает действие соединительных элементов, а фаза «операция» – нервно-импульсных элементов двухпроцессного механизма функционирования мозга. |
|
||
|
Главная | В избранное | Наш E-MAIL | Прислать материал | Нашёл ошибку | Наверх |
||||
|
|
||||