|
||||
|
|
Глава VII ПРИЗНАКИ НЕКОТОРЫЕ НЕЙРОАНАТОМИЧЕСКИЕ ПРОБЛЕМЫ ВОСПРИЯТИЯ К числу главных факторов, препятствующих пониманию неврологии процесса восприятия, относится одно из тех противоречий, один из тех парадоксов, с которыми мы неоднократно встретимся в этих главах, пытаясь связать функции нервной системы с психологическими процессами. В данном случае парадокс касается неироанатомическои организации системы входа и тех нарушений в поведении, которые возникают в результате разрушения этой организации (рис. VII-1). 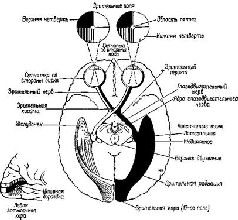
В сущности, анатомическая организация систем- входа является единственной организацией, в которой сохраняется топологическое соответствие между рецепторной поверхностью и корой головного мозга. Это означает, что расположение точек на сетчатке будет проецироваться на кору в виде сходного расположения точек. Таким образом могут быть построены карты ретино-кортикального соответствия. Такие карты строятся на основе данных, полученных либо методом дегенерации нервных волокон после небольших разрушений в рецепторных системах (Browner, 1934; Browner and Zeeman, 1926), либо в результате точечной электрической или световой стимуляции сетчатки при одновременной регистрации электрических ответов от участков ‹коры, в которых они могут быть вызваны (Talbot and Marshall, 1941). Подобные карты были пелучены для кожной чувствительности. По этому же принципу устроен и кохлеарно-кортикальный слуховой механизм (Rose and Woolsey, 1949). Такая рецепторно-кортикальная организация не сопровождается прямым однозначным соответствием посредством единой нервной нити. Скорее всего, столь же сложная организация, которую мы уже анализировали на уровне сетчатки, повторяется как на каждом уровне механизма рецепторной системы, так и в коре. Фактически наблюдается общее сокращение числа клеток в ганглиозном слое сетчатки по сравнению с рецепторным (у человека это уменьшение оценивается примерно как 100 или 50 к одному). При переходе от слоя ганглиозных клеток к латеральному коленчатому телу таламуса число зрительных элементов, по существу, остается постоянным. При переходе от таламуса к коре возникает обратная картина: одиночная .клетка латерального коленчатого тела может контактировать с 5000 кортикальных нейронов каждый из которых находится в контакте еще примерно с 4000 других нейронов через их дендритные связи. Благодаря такому механизму тормозных взаимодействий при стимуляции у обезьяны двух точек сетчатки в фовеа, несмотря на некоторое перекрытие зон возбуждения, возникает их раздельное восприятие, причем таким образом, что двум угловым минутам на сетчатке соответствует расстояние в 1 мм на поверхности коры (Talbot and Mar-chall, 1941, p. 134). Казалось бы, что такое строение нервной системы обеспечивает проекцию некоторого «образа» от рецеп-торпой поверхности на поверхность коры во многом так же, как проецируется изображение в кинокамере. Парадокс проявляется, когда в результате болезни или хирургической операции повреждаются системы входа. Правда, как и ожидалось, в некоторых случаях может быть обнаружено выпадение части поля зрения (скотома) в месте, которое может быть предсказано в соответствии с анатомической организацией мозга (рис. VII-2). 
Однако при сохранении хотя бы малой части входного механизма это выпадение часто не замечается даже в том случае, если глаза удерживаются в неподвижности. При этом распознавание структуры сохраняется и во многих отношениях не отличается от нормального. Люди с громадными скотомами либо совсем не знают о них, либо могут быстро научиться легко их игнорировать. Животное, у которого было удалено или разрушено от 80 до 98% нервных связей входного механизма, способно решать задачи, требующие различения структур, отличающихся только деталями. Лешли (1929) удалял у крыс 80-90% стриарной коры, не нарушая их способность различать зрительные структуры. Р. Галамбос перерезал у кошек 98% волокон зрительного тракта, и животные еще могли хорошо выполнять задания, требовавшие различения весьма сходных фигур (Galam-bos, Norton and Frommer, 1967). Недавно Чау (1970), также экспериментируя с кошками, разъединял более 3/4 волокон зрительного тракта и удалял более 3/4 зрительной коры; в таких условиях едва ли оставалась неповрежденной любая из проекционных систем, характеризующихся точечным («точка в точку») представительством. Хотя зрительное различение структур после такого значительного разрушения вначале оказалось нарушенным, животное снова научилось выполнять задание, на что ему потребовалось примерно такое же число проб, как и до операции. В моих экспериментах, как связанных с клинической нейрохирургией, так и проведенных в лаборатории (например, Wilson and Mishkin, 1959), удаление мозговой ткани было локальным и ограничивалось корой, так что оно не вызывало массивного разрушения белого вещества, благодаря чему у пациента или подопытного животного сохранялась в значительной мере способность к восприятию. После временно возникающей скотомы, длящейся несколько недель, можно было обнаружить лишь очень незначительные нарушения зрительной функции. Как уже отмечалось, исследователи пытались использовать и многие другие методы для разрушения предполагаемой ими организации входных систем мозга, но безуспешно: Р. Сперри и его группа (1955) хирургически производили перекрестное рассечение области, воспринимающей сенсорные сигналы, и даже помещали полоски слюды в места рассечения, чтобы электрически изолировать маленькие квадратики ткани друг от друга. Лешли, Чау и Семмез (1951) пытались накоротко замкнуть электрическую активность мозга, накладывая полоски золотой фольги на воспринимающие поверхности. Я создавал в воспринимающей области коры множество эпилептогенных очагов посредством инъекции микроскопического количества алюминиевой пасты (Kraft, Obrist and Pribram, 1960; Stamm and Pribram, 1961; Stamm and Warren, 1961). Хотя такое множество очагов заметно задерживало научение различению зрительных структур, оно не препятствовало его проявлению, если научение уже закончилось или происходило после того, как были произведены многочисленные разрушения. Эти результаты показывают, что эффект входного сенсорного воздействия на мозговую ткань, получаемая на входе информация распределяются по всей входной системе. Запись электрической активности мозга также свидетельствует о том, что в мозгу происходит распределение информации. Э. Р. Джон (John, Herrington and Sutton, 1967), например, при изучении зрительной системы использовал технику «меченых стимулов», предъявляя кошкам стимулы, которые различались не только своей геометрической структурой, но и частотой образующих эти стимулы мельканий. Эта различная частота мельканий отражалась на электрической активности мозга в ответ на предъявление стимула (или, если это мелькание было очень быстрым, то возникала субгармоника этой частоты). Таким образом, кодируемое различие частоты может оставить «след» в мозгу. Эта техника дала ряд интересных результатов, но для нас здесь важнее всего то, что тщательный анализ меченых волновых форм (вычисление позволяет обнаружить различие в волновых формах, возникающих в различных участках мозга) показывает, что одинаковые меченые волновые формы возникают во многих структурах мозга одновременно. 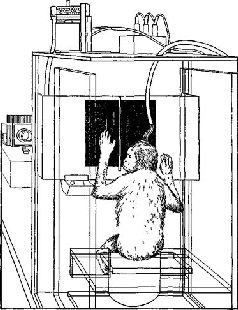

Однако другая группа экспериментов, проведенных в моей лаборатории (Pribram, Spinelli and Kamback, 1967, рис. VII-З, VII-4), показывает, что если научение уже имело место, то информация в мозгу не распределяется по всей системе. Были использованы очень маленькие электроды. Обезьяны были обучены по-разному реагировать на различные геометрические стимулы. В отличие от экспериментов Джона стимулы освещались очень короткой вспышкой света В зрительной коре было обнаружено несколько различных типов волновых форм электрической активности. Один тип был получен, когда форма волны определялась относительно момента предъявления стимула. Этот тип вызванного ответа обнаружил четкую зависимость от характеристики стимула. Два других типа были получены, когда форма волны определялась по отношению к началу ответа животного. Один тип волновой формы зависел от того, получила обезьяна подкрепляющий пищевой шарик за правильный ответ или не получила, когда ее ответ был ошибочным Другой тип волновой формы возникал непосредственно перед двигательной реакцией животного. Эта форма волны коррелировала с определенным типом двигательного ответа (нажатием на правую или левую панель); она следовала за стимулом, но не зависела от него и от полученного подкрепления. Важным для нас является тот факт, что все эти типы характерных волновых форм появляются локально в зрительной коре. Один тип волновой формы записан от одних электродов, другой от других. Распределение этих волновых форм происходило не в соответствии с какой-либо структурой возбуждения. Однако со дня на день, от недели к неделе наблюдалось полное постоянство в записях, получаемых от каждого отдельного электрода. Какой бы процесс кодирования ни происходил, он стабилизировался по мере того, как проводилась запись. Результаты этих экспериментов не согласуются с представлением о том, что образ, подобно фотографии, проецируется на поверхность коры. Они показывают, что каждая сенсорная система функционирует, имея большой резерв. Поскольку системы с разрушением и без него обнаруживают лишь незначительные различия в поведении, этот резерв, должно быть, распределен по всей системе – сохраняемая в ней информация, необходимая для осуществления различения, дублируется одновременно многими ее участками. Таким образом, вполне вероятно, что задержка в научении, возникающая при наличии эпилептогенных очагов, вызванных наложением на поверхность мозга алюминиевой пасты, обусловлена помехами, которые вносятся этими очагами в процесс дублирования хранения информации (рис. VII-5). 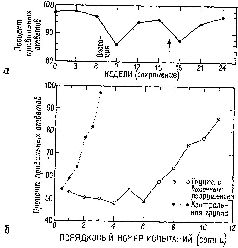
Вопросы, поднятые этими наблюдениями, следует сопоставить с другим рядом проблем: каким образом предметы кажутся нам достаточно стабильными, так что мы можем узнавать их независимо от угла зрения и расстояния до предмета? Каким образом мы узнаем предмет, несмотря на то, что светом, идущим от предмета, возбуждается только часть сетчатки, а следовательно, и мозга? Константность восприятия предметов и величины существует у детей уже в возрасте нескольких недель. Таким образом, простое объяснение феномена константности восприятия в терминах научения ставится под сомнение. Какого же рода механизм мог бы обеспечить одновременно существующую гибкость восприятия и константность узнавания? 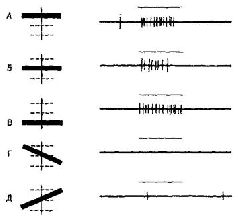
Как факты восприятия структур при наличии скотомы, так и факты константности восприятия говорят о том, что существует, должно быть, какой-то эффективный нейрологический механизм пространственного распределения информации на входе мозга. Для объяснения фактов восприятия недостаточно изоморфизма, простого («точка в точку») соответствия между стимулом и картиной возбуждения в коре, обусловленной анатомией системы. Когда в результате удаления коры происходит выпадение 80% зрительного поля, узнавание осуществляется благодаря оставшейся части зрительного поля; когда в зрительной коре производится множество разрушений, то сохранившиеся части функционируют столь успешно, что при различении испыты-ваются лишь незначительные трудности; рассматриваем ли мы предмет той или другой частью сетчатки, под тем или иным углом зрения, мы всегда можем узнать его. Восприятие отличается от обычного фотографического изображения – оторвите 98 или даже 80% У фотографии и попытайтесь идентифицировать ее: в большинстве случаев это невозможно. ПРОЦЕСС ВЫДЕЛЕНИЯ ПРИЗНАКОВ Один из способов решить эту дилемму – это отказаться от представления о том, что нейронный механизм входа является в каком-либо отношении устройством, формирующим образ, предположить, что узнавание структур есть результат операции выделения признаков, имеющей место на самом входе и возникающей с помощью отдельных нейронов или их небольших групп, и что восприятие зрительных структур осуществляется посредством иерархической системы таких «детекторов признаков». В пользу такого представления говорит то, что внутри частей входных каналов происходит конвергенция сигналов. Можно получить карты зрительных рецептивных полей для нейронов, находящихся на разных уровнях входной системы. В коре мозга было обнаружено два типа таких нейронов. Примерно у половины из них эти карты имеют форму более или менее округлых полей, аналогичных рецептивным полям ганг-лиозных клеток сетчатки, но в отличие от последних не имеющие четко выраженной окружающей зоны (Jung, 1961; Spinelli, Pribram and Bridgeman, 1971). Другая половина кортикальных нейронов со значительно большей вероятностью активируется входными системами, имеющими не округлые, а удлиненные формы. Чтобы вызвать максимальный эффект, эти удлиненные формы должны быть определенным образом ориентированы в отношении глаза, на который они воздействуют; разные нейроны отвечают на различную длину и ориентацию линий, границ и даже углов (Hubel and Wiesel, 1962; рис. VII-6). Известное объяснение этих наблюдений состоит в том, что нейроны, чувствительные к воздействию линии или границы, активируются только тогда, когда возбуждается большая часть или все линейное множество нейронов, чувствительных к воздействию таких пространственных признаков (рис. VII-7) и связанных с рецепторами линии и края. Высказано предположение, что благодаря дальнейшей конвергенции активность детекторов линии и края может комбинироваться и создавать клетки, реагирующие на более сложные структуры. Это предположение подтверждается тем, что найдены нейроны, особенно чувствительные к таким сложным структурам, как углы и короткие участки линий. Таким образом, объяснение восприятия на основе механизма анализа признаков становится крайне соблазнительным. Электрофизиологический анализ рецептивных полей клеток внутри вход-пых каналов показывает, что выделение признака в какой-то степени происходит внутри этих каналов. Далее, есть основание предполагать, что выделение основных признаков, таких, как цвет (De Valois, 1960; De Valois and Jacobs, 1968), контур и направление, осуществляется врожденными, встроенными в организм детекторами. Этот тип механизма, осуществляющего детекцию основных признаков, дополняется механизмом, обеспечивающим выбор признака, который зависит от прошлого опыта организма. Возникает вопрос, изменяет ли прошлый опыт сами эти детекторы признаков или в результате этого опыта в деятельность вовлекаются дополнительные нейроны, вызывающие изменения в нейронной популяции, изучаемой экспериментатором. Однако очевидно, что в зрелом организме процесс выделения признаков не может быть ограничен работой встроенных в организм детекторов. Должен происходить также анализ признаков посредством нейронов памяти. Поэтому обратимся теперь к логике той архитектоники нейронов, которая присуща анализаторам признаков, чтобы определить, что можно и что нельзя объяснить с этих позиций в самом восприятии. СТРУКТУРА АНАЛИЗАТОРОВ ПРИЗНАКОВ Софистическое утверждение Г. Вернера (1971), основанное отчасти на его собственных обширных исследованиях и работах В. Маунткасла (1957), Д. Хьюбела и Т. Визела и отчасти на концепции Т-О-Т-Е, относится к процессу анализа признаков в соместетической чувствительности и может быть с небольшими изменениями отнесено также и к другим входным системам. В коре мозга он различает базальную структуру – колонку, каждый нейрон которой обладает рецептивным полем, образующим единицу представительства входа. Эти нейроны имеют, как правило, идентичные или по крайней мере сходные рецептивные поля и, таким образом, образуют одинаковый уровень представительства. Эти колонки в свою очередь образуют более сложные структуры благодаря нейронам с дирекционпой чувствительностью, которые выполняют функцию указателей для согласования деятельности колонок (рис. VII-7).  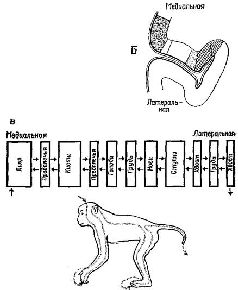
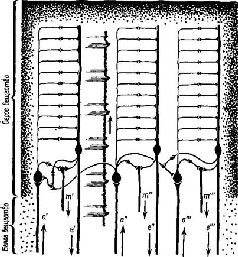
Эти указатели в зависимости от того, на какой ответ они настроены, определяют тот или иной тип взаимоотношений в электрической активности колонок; если имеются нейроны-указатели, которые чувствительны более чем к одной ориентации сигнала, то группы колонок оказываются связанными таким образом, что формируют «кольцевые структуры». Вернер сравнивает такую структуру коры с последовательным изложением инструкций, из которых составляются программы для электронно-вычислительных машин. Каждый лист программы содержит пункты, которые указывают на переходы к другим листам. Таким образом, посредством такого простого устройства могут быть запрограммированы сложные взаимодействия, последовательность операций. В терминах концепции Т-О-Т-Е наличие у колонок коры рецептивных полей со статическими свойствами создает условия для осуществления фазы проб в работе сервомеханизма; свойства колонок, свидетельствующие об их чувствительности к направлениям, составляют основу операционной фазы и фазы выхода. Сшшелли (1970) фактически составил программу (названную «Оккам»), моделирующую работу анализатора признаков, построенного па оспове такой кортикальной структуры. Предъявляя структуры нервных импульсов или волновых форм, можно настроить эту программу таким образом, чтобы она реагировала при повторении волн определенной формы (рис. VII-8). Несколько упрощепно точка зрения Вернера и Спинелли на анализатор признаков заключается в следующем: колонка коры состоит из входных и операционных нейронов, интернейронов и тестовых клеток. Входной сигнал, доходящий до нейронной единицы колонки, которая имеет свое рецептивное поле, распределяется по интернейронам. Последние в свою очередь связаны с операционными нейронами. Интерпейроны способны к настройке, то есть они адаптируются, обнаруживают привыкание и обладают памятью. Таким образом, каждый интернейрон действует как сумматор в электронно-вычислительной машине, который усредняет динамические структуры, вызванные поступающими на вход сигналами. Только при повторении таких динамических структур возбуждения в результате суммации возникает устойчивая структура, неповторяющиеся динамические структуры просто повышают нулевую линию и среднюю величину выходного сигнала. Таким образом, операционный нейрон, чувствительный только к динамическим структурам возбуждения, активируется лишь при повторении структуры входных сигналов. В целом процесс достигает своей завершенности благодаря посылке выходного сигнала от операционного нейрона к входному через тестирующий нейрон, который сличает структуру нейронной активности, вызванную входным сигналом, со структурой возбуждения, возникающей в операционных нейронах. Когда обе динамические структуры совпадают, тестирующая клетка продуцирует выходной сигнал, в противном случае при их несовпадении продолжается процесс настройки. Таким образом, каждая кортикальная колонка создает энгр:шму в соответствии со своей специфической чувствительностью к данной структуре нейронной активности, которая выступает в виде «описания» либо последовательности межспайковых интервалов, либо в волновой форме, которая являет собой огибающую узора спайковой нейронной активности. 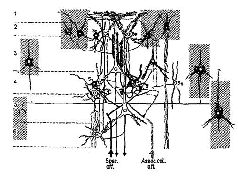
Считается, что каждая колонка связана с другими через горизонтальные клетки и их базальные дендриты, которые ответственны за тормозные взаимодействия (рис. VII-9). Всякий раз, когда эти горизонтальные клетки активируются несимметрично как это происходит, когда они возбуждаются через входы с ди-рекционной чувствительностью, создаваемые при этом временные структуры отдельных колонок образуют единую структуру. Следовательно, эти обширные структуры, зависящие скорее от гиперполяризации, чем от проведения нервных импульсов, обусловлены микроструктурой соединений. -Они и создают временно существующие состояния нейронов. Но более подробно мы остановимся на этом в следующей главе. Сейчас мы располагаем надежным доказательством того, что так называемые ассоциативные зоны коры осуществляют некоторый контроль над входными системами, во многих отношениях подобный тому, который имеет место при изменении кривизны хрусталика глаза. Такая функция контроля могла бы менять число (и, по-видимому, сложность) кортикальных колонок, которые могут участвовать в создании такой динамической структуры (гл. XVII). Таким образом, можно считать, что входные системы представляют собой некий фильтр для входных сигналов, экран, непрерывно настраивающийся на входные воздействия. Одной из характерных особенностей этого фильтра является то, что он создает самоадаптирующуюся систему, параметры адаптации которой находятся под контролем его собственной прошлой истории и тех операций, которые были осуществлены в нем посредством других нейронных механизмов. Другой особенностью фильтра как системы, которая обусловлена взаимосвязями между логическими элементами, является его способность к конструированию Образа. Но здесь мы забегаем вперед. Рассмотрим сначала проблему, почему необходим процесс формирования Образа вообще и в чем состоит ограниченность концепции анализа признаков при объяснении восприятия. ГРАНИЦЫ ПРИМЕНЕНИЯ ГИПОТЕЗЫ АНАЛИЗА ПРИЗНАКОВ Почему же неправильно рассматривать перцептивные процессы как безобразный анализ признаков? По трем причинам. Во-первых, анализируемые признаки не являются какими-то особыми, отличными от других. Во-вторых, богатство феномена восприятия не объясняется теми детекторами признаков, которые открыты. И в-третьих, изменения, которые имеют место в системах входа в ходе развития организма, позволяют различать эффекты, возникающие в детекторах признаков, изученные с помощью микроэлектродов, и эффекты, продуцируемые в системе, анализирующей эти признаки, что было изучено в поведенческих опытах с различением сигналов. Рассмотрим прежде всего проблему различимости признаков. И. Рок (1970) со всей проницательностью сформулировал эту проблему следующим образом: «… Те, кто утверждал бы, что ответ нейронных детекторов может объяснить, почему вещи выглядят такими, какими они являются на самом деле, возможно, не знают тех фактов, касающихся восприятия, на которые гештальтисгы обращали особое внимание. Например, известно, что воспринимаемая нами величина, не просто зависит от величины образа на сетчатке, воспринимаемая форма не является непосредственной функцией формы соответствующего изображения, а воспринимаемая скорость – непосредственной функцией скорости перемещения изображения на сетчатке и т. д. Напротив, несмотря на значительные изменения находящегося в отдалении от нас стимула, возникающие при изменении позиции наблюдателя, предметный мир сохраняет удивительную константность. Эти факты нельзя отрицать, и их нельзя оставить без внимания на том основании, что вещи в этих изменяющихся условиях не выглядят константными, а мы знаем, что они константны. Против этой формулировки имеются достаточно веские возражения. Рассмотрим детально некоторые факты, касающиеся восприятия движения. Начнем с того, что существует константность положения, а именно-известно, что, когда мы перемещаемся или двигаем глазами, перемещается и образ всей картины на сетчатке, но сама картина не кажется движущейся. Случается также обратное, а именно неподвижный образ на сетчатке вызывает восприятие движения при движении наших глаз или головы, как это наблюдается при слежении глазами за движущимся объектом или при рассматривании последовательного образа во время движения тлаз. Известно, далее, стробоскопическое движение, когда сменяющиеся неподвижные изображения вызывают ощущение движения. Известен также феномен индуцированного движения предмета, изображение которого на сетчатке неподвижно. Этот феномен возникает вследствие перемещения окружающих объектов. Так, например, Луна кажется движущейся на фоне перемещающихся облаков. Можно принять гипотезу, что действительной сенсорной основой восприятия движения является стимуляция детекторов движения, но тогда необходимо предположить, что значимость такой стимуляции определяется другой информацией. Например, можно сказать, что движение вообще не воспринимается, если данным о смещении изображения на сетчатке противостоит информация о том, что оно возникает в результате собственного произвольного движения организма, как это предположил фон Хольст, -а до него Гельмгольц. Или можно считать, что в случаях индуцированного движения детекторы предоставляют решающую информацию о том, что что-то движется, но по неизвестным еще причинам движение приписывается неподвижному объекту. Откровенно говоря, я не считаю это достаточно убедительным. Это не объясняет, почему я вижу свой последовательный образ движущимся, когда я двигаю глазами даже в совсем темной комнате, где не видно неподвижных предметов, изображения которых могли бы перемещаться на сетчатке. В случае же стробоскопического движения, если наши глаза удерживаются в стационарном положении, то источником стимуляции детекторов движения мог бы быть только возникающий в этих условиях нистагм глаза. Но это не могло бы объяснить движения, которое возникает между двумя раздельно освещаемыми объектами. Эта гипотеза не может объяснить и индуцированного движения, потому что предмет, который вызывает ощущение движения, может двигаться так медленно, что его движение будет ниже порога, в то время как только оно и воспринимается. Далее, очевидно, что в данном случае основой восприятия движения является изменение локализации предмета, который кажется движущимся относительно местонахождения предмета, который вызывает это движение, а не стимуляция детекторов перемещения изображения. Рассмотрим теперь следующую проблему – проблему восприятия ориентации предмета. Является ли основой воспринимаемой ориентации линии ориентация ее изображения на сетчатке? Очевидно, нет, поскольку при повороте головы вертикальные и горизонтальные линии на стене продолжают оставаться для нас вертикальными и горизонтальными. Здесь мы «нова сталкиваемся с константностью, в данном случае с константностью восприятия направления, несмотря на изменения в ориентации изображений на сетчатке. Это происходит в значительной степени даже тогда, когда мы рассматриваем в темноте одну светящуюся линию, наклонив или откинув голову в другую сторону. Следовательно, обоснованно заключение, что, прежде чем сделать вывод о значении для восприятия ориентации данного сетчаточного образа, необходимо принять во внимание информацию о собственной позиции наблюдателя. Далее, как бы мы поняли открытие, сделанное Хьюбелом и Визелом, о существовании в зрительной коре детекторов ориентации? Очевидно, что они не могут быть знаками ориентации объектов в окружающей среде. Можно согласиться, что они являются знаками, указывающими на ориентацию объектов относительно организма, другими словами, детекторами эгоцентрической ориентации, и что они создают условия, необходимые для того, чтобы организм мог определить ориентацию объектов в окружающей среде, когда он дополнительно получает информацию и об ориентации самого ' наблюдателя. Например, можно сказать, что, когда изображение линии на сетчатке будет иметь вертикальное положение, оно всегда будет казаться эгоцентрически вертикальным, то есть параллельным продольной оси головы, и линия будет восприниматься вертикальной в поле зрения наблюдателя, если он занимает вертикальное положение, и горизонтальной, если наблюдатель повернут на 90°. Хотя такое объяснение кажется мне правдоподобным, оно вызывает серьезные трудности из-за существования некоторых дополнительных фактов. Если наблюдатель сохраняет вертикальное положение, то вертикальная линия, воспринимаемая им на фоне изображения комнаты, при наклоне последнего будет выглядеть наклоненной в направлении, противоположном наклону изображения комнаты. Таким образом, в данном случае вертикальное изображение на сетчатке не будет более соответствовать вертикально расположенному предмету, несмотря на то, что наблюдатель находится в вертикальном положении. Более того, линия не будет уже казаться эгоцентрически вертикальной линией. К этим фактам можно было бы добавить данные о том, что наблюдатели будут адаптироваться к повернутым с помощью призматических стекол изображениям, так что вертикальное изображение на сетчатке не будет больше ни знаком вертикального объекта, когда наблюдатель находится в вертикальном положении, ни фактически знаком того, что объект ориентирован в вертикальном направлении относительно наблюдателя. Имеют ли какое-либо отношение детекторы ориентации в коре к восприятию формы? Можно предположить, что восприятие формы сводится к восприятию ориентации линий, из которых строится форма. Таким образом, предмет воспринимается как квадрат тогда, когда одновременно стимулируются два детектора вертикальных и два детектора горизонтальных линий. Хорошо известно, что фигуры выглядят весьма по-разному, когда меняется их ориентация. На первый взгляд сам этот факт может казаться доказательством того, что детекторам ориентации принадлежит существенная роль в восприятии формы, потому что в этом процессе принимают; участие совершенно различные волокна. Можно сказать, что это является основой для понимания явления изменения внешнего вида предмета. Однако в действительности, вообще говоря, объяснением изменения внешнего вида предмета служит не изменение ориентации изображения на сетчатке. В проведенных мною и моими сотрудниками экспериментах неоднократно-было показано, что, если положение фигуры в отношении ее окружения не меняется, но она рассматривается наблюдателем в наклонной позе, фигура не выглядит измененной, или, выражаясь более точно, распознавание не ухудшается. Могло ли бы это быть, если за это явление были бы ответственны совершенно различные детекторы? Напротив, если фигура наклонена так же, как и наблюдатель, и ориентация изображения на сетчатке, таким образом, совсем не изменилась, фигура тем не менее выглядит совершенно иной и иоэтому часто не узнается. Можно провести простой опыт: создайте последовательный образ квадрата, держа голову в вертикальном положении, а затем, закрыв глаза и повернув голову на 45°, посмотрите на него, он будет подобен ромбу. Могло ли бы это быть, если за это явленяе были бы ответственны одни и те же детекторы? Однако эти факты приобретают смысл, если считать, что решающим-для ориентации является восприятие определенной части фигуры как верх, а другой – как низ и т. д. Если мы будем правильно оценивать одну и ту же часть фигуры в качестве верха, как мы это обычно делаем, приняв наклонное положение, то мы увидим, что форма предмета не меняется. Однако, если мы за верх фигуры принимаем другую ее часть, как это бывает при наклоне фигуры относительно окружения, то мы увидим, что ре форма очень изменилась. Этот процесс, по-видимому, не имеет ничего-общего со специфическими детекторами ориентации. Далее, роль этих детекторов, возможно, состоит в том, что они служат для различения ориентации одного образа по отношению к другому. В прошлом мы ошибочно считали, что такое различение осуществляется благодаря различной пространственной ориентации проекционных кортикальных «образов». Иными словами, нейронные детекторы, вероятно, являются механизмами, посредством которых воспринимается определенная информация о стимуле, в данном случае информация о том, что сохраняется или изменяется пространственная ориентация. Конечно, такая информация необходима для восприятия в качестве исходной, но ответ волокон детекторов никоим образом нельзя рассматривать как нейронные корреляты единиц восприятия» (1970, р. 2-5). 
В остальной части статьи Рока анализируется еще ряд интересных экспериментов, которые трудно объяснить с позиции детекторов признаков (см. также рис. VII-10, VII-11). 
Второй недостаток точки зрения, рассматривающей восприятие как безобразный процесс выделения признаков, состоит в том, что по сравнению с богатством явлений сознания процесс распознавания структуры, осуществляемый методом классификации, довольно беден деталями, которыми он оперирует. Механизм распознавания зрительной структуры, который осуществляется путем избирательного выделения признаков, может приводить к ошибкам. Каждый из нас ждал кого-нибудь, кто опаздывал на свидание, назначенное в многолюдном месте, и ошибочно принимал посторонних за ожидаемое лицо, так как они имели сходные с ним черты. Мы узнаем А, Л и а как букву А; мы хорошо знаем о различиях почерков. Восприятие, по-видимому, обеспечивает в одно и то же время не только богатство нашего опыта и «сильное предпочтение сохранять постоянную организацию», но также и тенденцию к «максимальному упрощению», как показали опыты гештальтпсихологов. Имеются данные, свидетельствующие о том, что богатство восприятия связано со сложностью конфигураций окружающей среды, которые существуют независимо от организма и возбуждают его рецепторные поверхности (см. Gibson, 1966), последние приводят в действие некоторый механизм мозга с соответствующей емкостью, позволяющей вместить это богатство. Данные, представленные Роком и другими, показывают, что только механизм выделения признаков обусловливает тенденцию к упрощению, создает схему отношений. Однако даже здесь начинает играть роль другой механизм – более сложный «познавательный» процесс. Третий недостаток объяснения фактов восприятия с позиции выделения признаков выступает в экспериментах, в которых новорожденные в течение различных периодов времени подвергались сенсорной депривации. Как уже говорилось в гл. VI и VIII, младенцы в возрасте нескольких недель обнаруживают способность к восприятию константности величины и формы. У животных способность к различению развивается также рано; при изучении мозга детенышей животных были обнаружены детекторы признаков, которые уже достигли своего полного развития. Однако при ограничении потока сенсорных воздействий эта простая корреляция между поведением и функциями мозга нарушается. Так, когда кошки и крысы содержатся в полной темноте, их способность различать яркость и контур остается неизмененной, но способность различать более сложные зрительные структуры значительно ухудшается. Хотя ориентация рецептивных полей кортикальных нейронов у таких животных еще различима, она имеет менее острую настройку, и утомление клеток развивается быстрее, чем у животных, содержащихся в нормальных условиях. Но когда сенсорная депривация ограничена депри-вацией одного глаза в течение определенного времени, как зрительная ориентация, так и способность к различению оказываются резко нарушенными, если животное пользуется только одним этим глазом. Это связано с тем, что стимуляция глаза, подвергнутого депривации, может привести в активное состояние лишь небольшое число нейронов, которые вряд ли могут сколько-нибудь избирательно отвечать на определенным образом ориентированную линию или движение. Эта реакция возникает, даже несмотря на то, что в коре присутствует обычный для нее комплект клеток с их нормальной реактивностью, как это было обнаружено при стимуляции не подвергавшегося депривации глаза (см. рис. VII-12). Таким образом, когда имеет место нарушение в привычном способе возбуждения, происходит какое-то подавление реактивности и это подавление переходит в нарушение функции исключительно в результате неупотребления нервных структур (как у животного, подвергшегося полной сенсорной депривации). Если монокулярное подавление реактивности коры объясняется некоторым изменением в связях, то ответственным за этот эффект подавления должен быть какой-то процесс, отличный от простого выделения признаков. 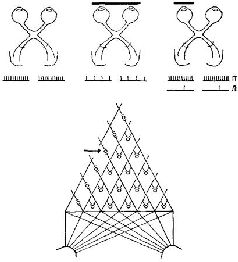
Л. Ганц в недавнем обзоре (1971) этих и других сходных данных доказал это. Основываясь на результатах экспериментов с полной депривацией животных, он различает два типа заданий. Ориентация по яркости и контуру требует только, чтобы организм отвечал на языке реакций, свойственных большинству его детекторов признаков. Различение более сложных структур, например двух треугольников, один из которых перевернут, не может осуществляться на этой основе. Мы нуждаемся в каком-то дополнительном, более гибком мозговом механизме, который Ганц называет «избирательным вниманием». Этот механизм активно подавляет доминирующую реакцию детекторов признаков, реакцию, которая, в сущности, не имеет отношения к адекватному решению задачи на различение. Следовательно, мы снова сталкиваемся с необходимостью иметь дополнительный механизм, помимо выделения признаков. Поэтому мы снова приходим к мысли о той, что, возможно, системы входа организованы таким образом, что нейронные сигналы координируются с каким-то психологическим процессом формирования образов. Проблема состоит в том, чтобы определить нейро-логическую природу этой организации. На основе рассмотренных в начале этой главы данных становится очевидно, что эта организация не может быть просто мозаикой точек, которые генерируются в рецепторах и без изменения проводятся до коры, где они формируют образ, подобно фотографии. Столь же очевидно, что следует принять во внимание наличие нейронов, чувствительных к линиям, и других детекторов признаков, которые создают в мозгу нейронную основу для их анализа. РЕЗЮМЕ Почти полное удаление мозговой ткани независимо от ео л№-кализации в пределах систем входа оказывает лишь незначительное влияние на процесс распознавания. Следовательно, логика работы нейронов, свойственная любому конкретному восприятию, распространяется, по-видимому, на всю систему. Были получены также прямые данные, касающиеся распределения памяти в мозгу. Логику этой памяти создают как встроенные детекторы элементарных признаков (таких, как линии и углы), так и анализаторы для более сложных преобразований особых признаков (таких, как волновые формы), которые перестраиваются в ходе жизненного опыта организма. |
|
||
|
Главная | В избранное | Наш E-MAIL | Прислать материал | Нашёл ошибку | Наверх |
||||
|
|
||||