|
||||
|
|
Глава III ЗАТУХАНИЕ АКТИВНОСТИ В НЕРВНОЙ СИСТЕМЕ И ТОРМОЗНЫЕ ВЗАИМОДЕЙСТВИЯ ОРИЕНТИРОВОЧНАЯ РЕАКЦИЯ И ПРИВЫКАНИЕ В первой главе были проанализированы два основных класса переменных, характеризующих деятельность мозга. Взятый в качестве модели процесс воспроизведения речи показал возможности, открывающиеся благодаря модификации этих двух переменных. Во второй главе были детально рассмотрены данные, свидетельствующие о пластичности мозга и его способности к постоянным изменениям; однако мы еще не говорили о свойствах самого механизма, который делает возможным осуществление таких изменений. При игре на скрипке, флейте или органе участвуют те же самые основные переменные, что и в речевом общении, хотя сами инструменты различаются тем, как они воспроизводят звуки. Поэтому настоящая глава начинается с рассмотрения организации нервной системы, позволяющей ей быть аппаратом, создающим все богатство нашего опыта и поведения. Эта организация во многом зависит от относительно постоянных структур, образующихся в тех системах, посредством которых мозг приходит в соприкосновение со своим окружением. Благодаря своей доступности эти системы могут служить некоторыми частными моделями и, следовательно, подвести к исследованию более центральных процессов, анализ которых тебует иногда больших усилий. Поскольку довольно сложная организация имеет место уже при первоначальном контакте организма с его окружением, уместно начать наше исследование именно с этого процесса. Экспериментальные данные говорят о том, что в любой момент времени происходит отбор сенсорного возбуждения с помощью хранящейся в памяти информации о прошлом опыте. Этот процесс сравнения, в ходе которого выявляется, соответствует ли текущее возбуждение хранящимся в мозгу записям, направляет как внимание человека, так и его действия. Е. Н. Соколов (1960) провел следующий простой опыт, в котором обнаружилось одно из тех противоречий, которые порождают полезные парадоксы и указывают направление экспериментальным исследованиям в различных разделах нейропсихологии. Человеку предъявляется звук гудка. Сначала он вызывает ориентировочный рефлекс. Электрическая активность мозга характеризуется следующими чертами – активацией в латеральных отделах больших полушарий головного мозга (в виде увеличения доли низкоамплитудных и высокочастотных компонентов ЭЭГ) и гиперсинхронизацией в медиальных и базальных структурах мозга, на что указывает возрастание ритмичности и рост амплитуды волн среднечастотного диапазона. Кроме того, возникают другие локазатели ориентировочного рефлекса: увеличивается приток крови к голове за счет оттока от кончиков пальцев; меняется электрическое сопротивление кожи, частота сердцебиения и дыхания. Однако при многократном повторении звука гудка все эти реакции заметно уменьшаются. В таком случае говорят, что испытуемый обнаружил привыкание к стимулу. Однако исчезновение реакции на повторяющиеся звуки оказывается обманчивым. В действительности многие из компонентов этой реакции еще сохраняются, так как, если стимул слегка меняется (например, звук становится тише), все первоначально исчезнувшие реакции восстанавливаются. Конечно, это парадокс. До открытия этого явления Соколовым высказывалось предположение, что привыкание просто повышает порог нервной системы на входе. Результаты, полученные Соколовым, означают, что лицо, которое обнаружило привыкание, должно было сопоставить действующий звук с сохранившимся представлением о ранее воспринятых звуках – иначе почему бы снижение интенсивности звука снова вызвало полноценную ориентировочную реакцию? Соколов проверил свою интерпретацию, многократно предъявляя испытуемому тон определенной длительности. Затем он неожиданно предъявлял более короткий звуковой раздражитель. Сразу же после прекращения звука возникали ориентировочные реакции; таким образом, изменение ответов происходило при наступлении тишины. Эти реакции продолжались в течение всей «ожидаемой» длительности звука, а затем медленно исчезали. Мы все испытывали эту удивительную реакцию на неожиданное наступление тишины (рис. III-1). Я назвал это «феноменом Бовери». В течение многих лет на Третьей авеню в Нью-Йорке существовала надземная железная дорога, которая производила ужасающий шум; когда она была снесена, люди, жившие в домах вдоль железной дороги, периодически просыпались от отсутствия звуков во время сна и звонили в полицию, чтобы сообщить, что происходит нечто странное, хотя они не могли точно определить, что именно. Эти звонки раздавались в те моменты, когда раньше с грохотом проносились поезда. Появление чего-то странного было, конечно, связано с наступлением «внезапной тишины», которая заменила ожидаемый шум. 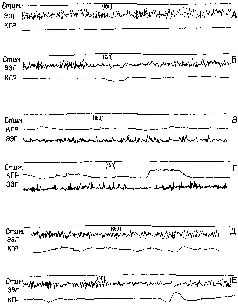
Множество данных свидетельствует о том, что ориентировочные реакции и привыкание длятся сравнительно долго и являются в высшей степени специфическими. Каждая часть мозга характеризуется своими электрическими сигналами и своей последовательностью развития электрических явлений при осуществлении ориентировочной реакции. Если возникает привыкание, то нарушение его (ориентировочная реакция) при всяком изменении ситуации продолжает возникать на протяжении шести месяцев; изменения в последовательности событий, в обстановке, в которой они происходят, или в интенсивности, в длительности или конфигурации самих событий будут вызывать восстановление ориентировочных реакций. Нейрофизиологии привыкания посвящено много исследований, и она вызвала много споров из-за различных экспериментальных подходов к решению этой проблемы. Для того чтобы стало возможным объяснение этого явления, необходимо уточнить некоторые факторы. Большинство нейрофизиологов называют привыканием любое постепенное ослабление ответа при повторном предъявлении того же самого стимула. Психологи же считают привыканием только те случаи, когда затухание ответа имеет место после нескольких проб и когда легко вызвать восстановление ориентировочной реакции. Недавно некоторые исследователи мозга, в том числе и я, приняли еще более строгий критерий Соколова: неожиданное уменьшение интенсивности, длительности стимула или его отсутствие должны вызывать восстановление ориентировочного рефлекса, если прежде этот рефлекс угасал в результате привыкания. Важно также место электродов при электрофизиологической записи. Так, в моей лаборатории мы обнаружили, что привыкание действительно характеризуется затуханием ответа на любом уровне зрительной системы обезьяны от сетчатки до коры головного мозга. Однако при разном расположении электродов в коре одни из них дают признаки ослабления, другие – увеличения ответа, а третьи не обнаруживают никаких изменений. Следовательно, при повторении стимула в коре возникает изменение структур возбуждения; растормаживание разрушает эту структуру. Если бы мы смешали записи от всех кортикальных отведений или сделали бы записи от черепа, а не через электроды, вживленные непосредственно в кору, мы не смогли бы обнаружить этот эффект, о чем действительно сообщают некоторые исследователи. Много работ было выполнено и на клеточном уровне. Была произведена запись от отдельных клеток спинного мозга и других отделов мозга. Во многих местах были обнаружены нервные клетки с затуханием ответа при повторении стимуляции. Такая реакция возникает даже у беспозвоночных, например у Aplesya Cali-fornica (Pinsker et al., 1970; Kupfermann et al., 1970; Castellucci et al., 1970). Однако можно ли у всех нейронов, у которых обнаружено привыкание, вызвать явления растормаживания? Недавно полученные Р. Томпсоном данные (Grove, Thompson, 1970) говорят о том, что можно различать по крайней мере три группы нейронов: нейроны, у которых реакция просто затухает; нейроны, реакция которых первоначально усиливается; и нейроны, характеризующиеся сначала усилением, а затем затуханием реакции. -Лишь у последней группы возможно растормаживание. Такие эксперименты до сих пор были проведены только на спинном мозге. Другие отчеты об экспериментах на структурах ствола мозга (верхнее двухолмие) свидетельствуют о том, что возможно, хотя и не обязательно, что одиночные нейроны с затухающими ответами в этой структуре могут обнаруживать растормаживание (Gerbrandt, Bures, Buresova, 1970). Эти данные, а также другая работа Томпсона (Thompson and Spenser, 1966) показывают, что привыкание связано с взаимодействием нескольких нейронов. Томпсон использовал препарат, принесший известность Шеррингтону: животное, у которого спинной мозг отделен от головного. Повторная стимуляция электрическими залпами входных волокон спинного мозга приводит к постепенному уменьшению рефлекса сгибания конечности (вызванного подобно коленному рефлексу у больного, подвергающегося врачебному осмотру). Этот препарат создавал идеальные условия для изучения места и физиологической природы тех изменений в спинном мозге, которые связаны с привыканием (рис. III-2). Ни входные, ни выходные нейроны не участвуют в изменении рефлекса и, следовательно, не ответственны за привыкание: решающее значение имеют интернейроны, связывающие нейроны входа и выхода. Во-вторых, вопреки ожиданию введение фармакологических веществ, обычно используемых нейрофизиологами при изучении пре- и постсингштических .эффектов, не оказывало никакого влияния; эти вещества не приводили к затуханию ответа, свидетельствующему о привыкании. Это значит, что привыкание не связано с изменениями медленных потенциалов соединительных контактов. Сходные результаты получил Соколов (Соколов, Пакула и Аракелов, 1970). Он обнаружил, что медленные потенциалы мембраны нейрона и генераторные потенциалы, записанные от его аксона, которые дают начало нервным импульсам, могут изменяться независимо друг от друга. Кроме того, полученные им результаты свидетельствуют о том, что механизм генерации нервных импульсов может обнаружить привыкание. 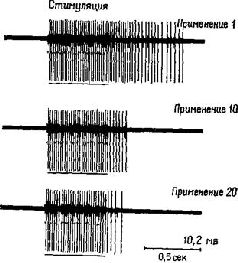
Таким образом, в некоторых случаях процессы затухания, лежащие в основе привыкания, по-видимому, связаны не с пре- и постсинаптическими изменениями, а с какими-то другими механизмами. Однако в других случаях были получены противоположные результаты. Иногда \по не всегда) этот процесс связан с уменьшением или увеличением притока медиаторных веществ, С. Шарплесс (1967, 1969) показал, что при стимуляции нейронов у позвоночных не возникает ни сенситизации, ни десенси-тизации их сияаптических мембран. Напротив, Э. Кендел (Castel-lucci et al., 1970; Kupfermann et al., 1970; Pinsker et al., 1970) нашел, что привыкание и его нарушение у беспозвоночных локализуются в пресинаптических окончаниях и предполагают участие эффективного механизма секреции возбуждающих веществ в синапсах. Стало быть, полученные данные не согласуются между собой. Необходимо найти ответ на целый ряд вопросов. Каким образом вообще механизм возникновения возбуждений может испытывать влияния, если не через посредство медленных потенциалов? Запускается ли внутри нейрона некоторый биохимический процесс и если да, то как (особенно если мы имеем дело с системой, которая обычно обнаруживает спонтанную активность и, следовательно, каким-то образом генерирует потенциалы)? Какова природа биохимических изменений и как они развиваются во времени? Соколов предполагает, что в этом механизме участвует РНК. Если он прав, то не связано ли привыкание с первыми этапами индукционного процесса или механизм привыкания совершенно независим от процесса постоянного хранения следов памяти? Возможно, что механизм постоянного хранения действует только при условии длительного и повторного привыкания. Если это так, то как это происходит? Наконец, каким образом изменение в структуре нервного импульса, вызываемого привыканием, действует на микроструктуру медленного потенциала на следующем синаптическом уровне? Чтобы ответить на некоторые из зтих вопросов, рассмотрим нейронную организацию сетчатки, этого маленького «кусочка мозга», который является гораздо более доступным для изучения, чем центральная нервная система. |
|
||
|
Главная | В избранное | Наш E-MAIL | Прислать материал | Нашёл ошибку | Наверх |
||||
|
|
||||